News Center
热点新闻
免疫亲和柱市集份额2023理解陈说免疫亲和柱运用手段肝素亲和柱道理免疫亲和柱能够反复
阐明驱动抗原识另表互相功用机造,是剖判抗体介导的爱惜功用的中央,也是合理打算免疫原的合节。现有表位解析工夫(如单克隆抗体咨询、深度突变扫描、酵母展现平台等)多依赖重组卵白或已知单克隆抗体,难以正在自然血清境况中整个解析多克隆抗体的繁复表位组合。现有法子难以检测构象表位(如线性表位工夫),且无法反应自然血清中多克隆抗体的的确互相功用。
氢氘换取质谱工夫(HDX-MS)通过监测卵白质主链酰胺氢的氢/氘换取率转化,可能绘造构象表位,并追踪抗体连接惹起的变构效应。液相色谱-质谱联用(LC-MS)仪器的连续提高,进一步拓展了HDX-MS正在解析更繁复、异质性卵白复合物方面的材干。先前咨询已表明,该法子可能同时定位多个单克隆抗体(mAbs)连接的表位,以至可解析从血清中富集的多克隆抗体(pAbs)夹杂物的表位特点2。为了拓展该工夫,本文通过整合固定化抗原与HDX-MS工夫,治理了守旧表位解析依赖人为重组体例、丧失自然构象讯息、无法解析多克隆抗体繁复靶向形式等瓶颈。其正在模仿的确境况下对表位举行定位,可认为疫苗开垦、熏染免疫机造咨询及调理性抗体筛选供应了更亲昵心理条款的解析用具。
本文用到的工夫流程如图1所示。作家将生物素化的抗原SEB通过链霉亲和素固定于磁珠表表,保存其native构象。将固定化的SEB与区别源泉的抗血清(抗SEB绵羊血清、抗SEB兔血清、native兔血清)孵育,随后彻底洗涤未连接抗体。将抗原-抗体复合物置于氘代缓冲液中举行氢氘换取反映,并通过变性、还原(此时抗原与磁珠解离)、酶切、-MS,监测未孵育血清与孵育血清后的SEB的HDX速度转化。
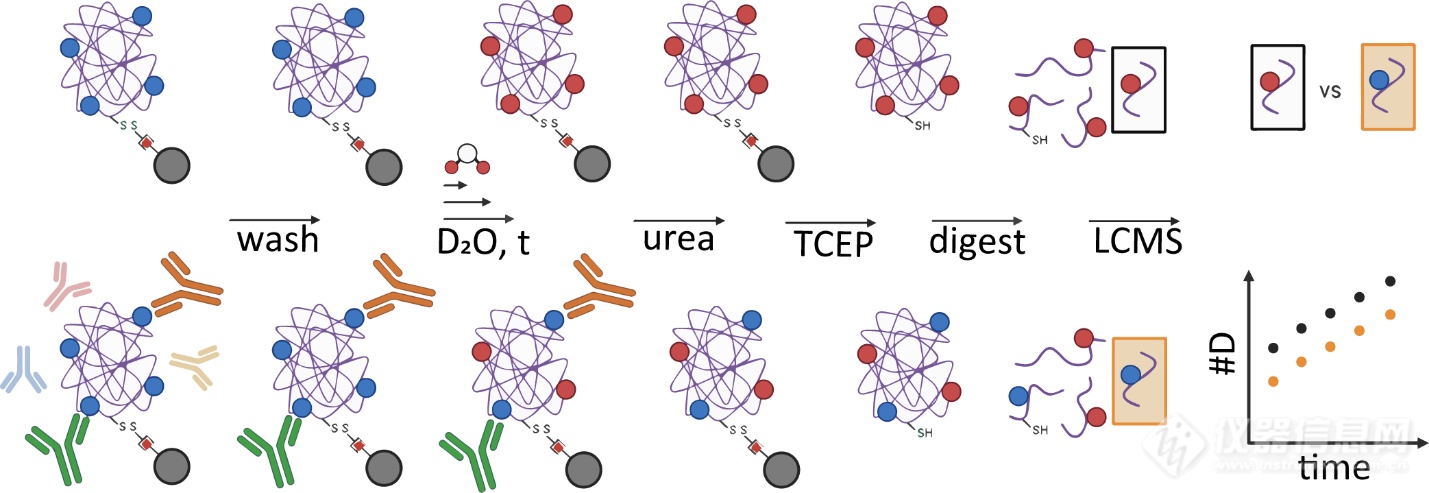
本文起初运用生物层干预工夫(BLI)测定mAbs与SEB的连接动力学参数(kon、koff)。以验证三种单抗(20B1、14G8、6D3)对SEB的连接亲和力,为后续HDX-MS咨询供应亲和性格根本。悉数mAbs均显示pM级高亲和力,但14G8的连接妥协离速度比其他mAbs慢约10倍(动力学图未正在此展现)。三种mAbs均与SEB高亲和力连接,但需当心14G8的动力学性格能够影响其功用。
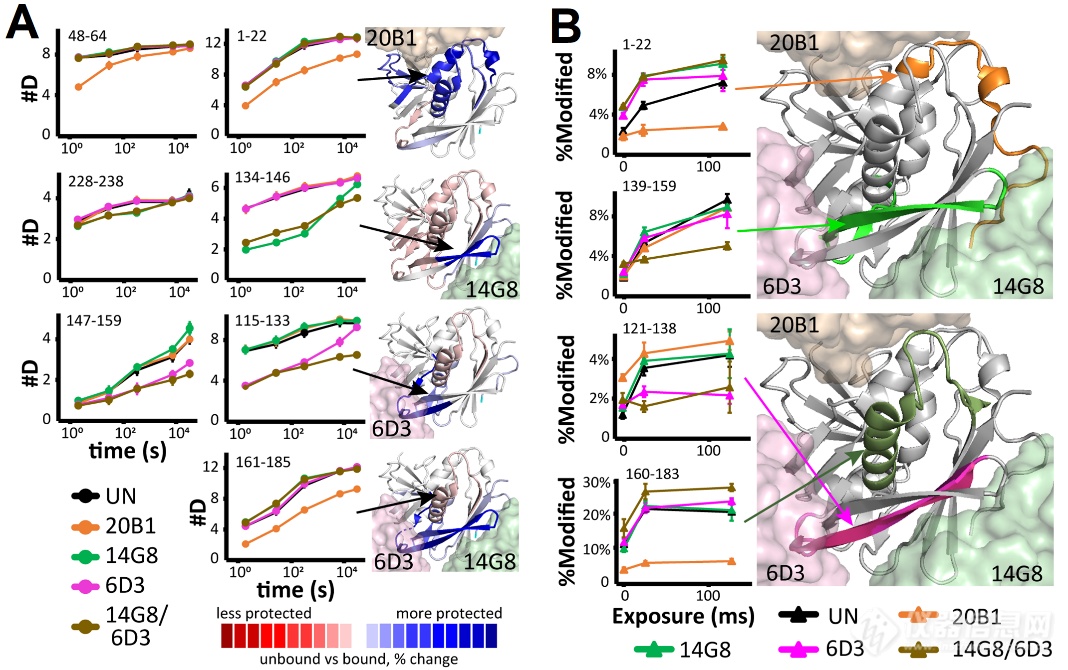
接着作家通过HDX-MS解析了mAbs与SEB的连接界面。作家检测了三种单抗20B1、14G8、6D3稀少与SEB孵育以及14G8/6D3夹杂与SEB孵育时的氘代转化景况。文中出现三个单抗的氘代低落区域与晶体布局报道的连接区域相似(图2A)。但14G8/6D3夹杂组显现了分表爱惜区域134-146(14G8界面)、115-133和147-159(6D3界面)。这注解14G8/6D3夹杂发生了协同爱惜效应,这能够反应其正在体内的协同爱惜机造。其它,作家用羟基自正在基X射线踪迹(XRF)验证连接界面,该工夫通过测定侧链溶剂可及性转化验证连接界面,能够增补HDX-MS结果。此中,监测到的20B1和6D3连接区域(氧化削减区域)与HDX-MS出现的连接区域基础相似(图2B)。但也存正在氧化上升的区域,如20B1的121-138区域、6D3的1-22区域,提示这些区域的侧链存正在连接惹起的变构效应。而14G8/6D3夹杂组显现139-159爱惜、160-183去爱惜局面,提示14G8/6D3夹杂发生特有连接效应,撑持协同爱惜机造。
接着,作家分辨解析了兔和羊血清中抗体靶向SEB的表位不同。如图4A,作家出现兔和羊血清中对SEB的137-146和198-208区域都拥有氘代低落趋向。区另表是,兔血清对36-43区域发生氘代爱惜,52-64区域氘代扩充。而羊血清对115-133发生氘代爱惜。这注解区别物种血清靶向SEB的表位存正在不同,能够与抗体库构成相合。
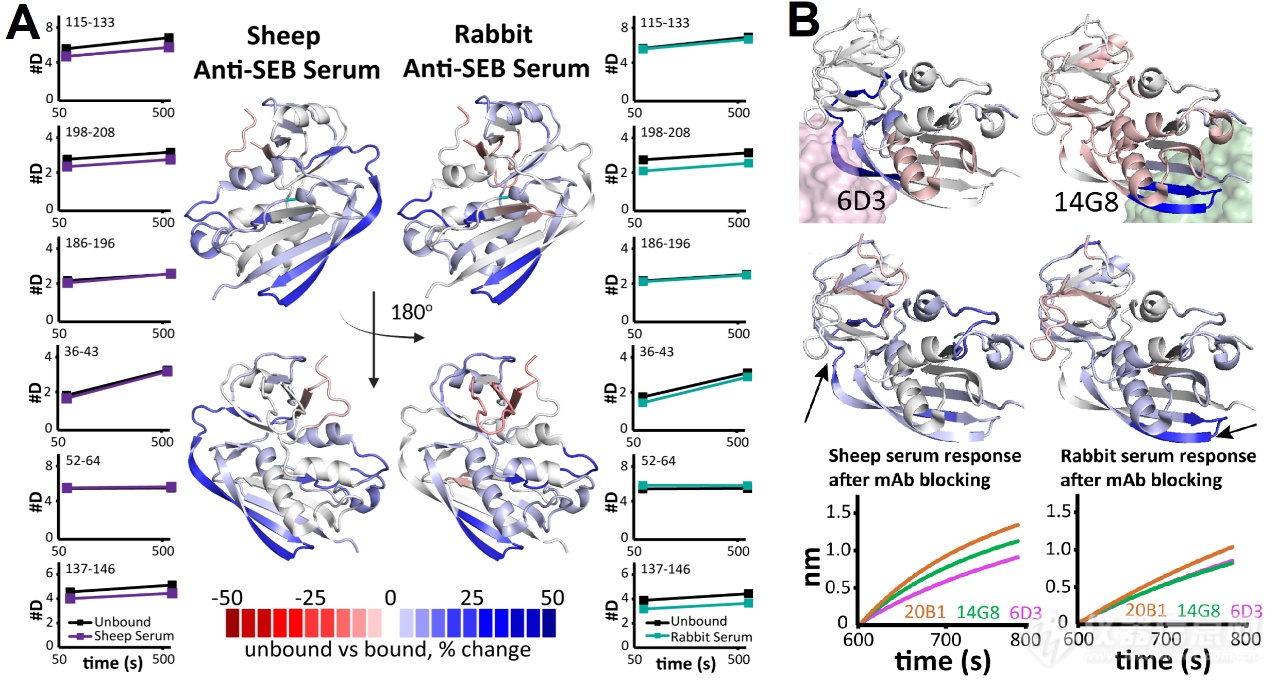
结果,为了验证通过多克隆固定化SEB HDX-MS法子确定的苛重表位,作家运用已知特异性的单克隆抗体(mAbs)举行了BLI逐鹿试验。起初,残基115-133(位于6D3的表位内)仅正在羊血清中显示爱惜,残基137-146(位于14G8的表位内)正在兔血清中爱惜更明显(图4B)。而事先用mAbs 6D3封锁的固定化SEB与羊血清的连接明显低落,而用14G8封锁后的固定化SEB兔血清的连接明显低落。这验证了固定化SEB HDX-MS出现的羊血清抗体苛重连接正在6D3对应的表位上,而兔血清抗体苛重连接正在14G8对应的表位上。


